

一般的な脊椎動物の組織は、細胞質、細胞間質液、脂質二重膜、そしてタンパク質といった屈折率(Refractive Index: RI)の異なる多様な構成要素から成る不均一な構造体である。光がこのような微小な不均一な媒体を通過する際、ミー散乱およびレイリー散乱と呼ばれる光の散乱現象が引き起こされる。ほとんどの生体組織において、散乱係数は吸収係数の10倍から1000倍に達するため、結果として組織は不透明となる。しかし、透明な魚類は、遺伝子レベルでの色素形成の阻害、光学的バリアを持たない特異な皮膚構造、そして筋肉組織の極めて精緻な光学的配置を組み合わせることで、光の散乱を極限まで抑制している。
特筆すべきは、この「光学的マジック」が生体の厳密なホメオスタシス(恒常性)に依存した動的な現象であるという点である。生体が極度の環境ストレスに晒されたり、死を迎えてエネルギー供給が途絶えたりした瞬間に、細胞内の秩序は崩壊し、透明な体は一気に乳白色へと白濁してしまう。この事実は、生体における透過性という特性が単なる静的な物理構造の産物ではなく、多大なエネルギーを消費して能動的に維持される現象であることを明確に示唆している。
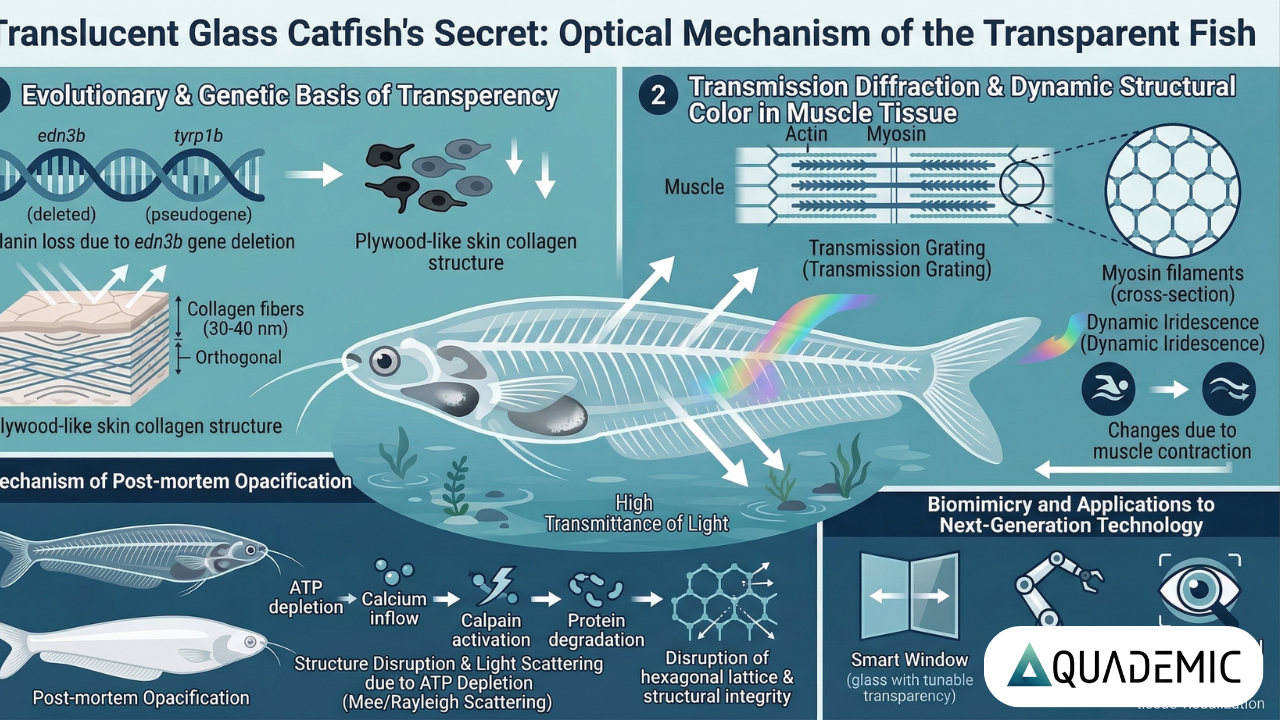
透過性の進化的および遺伝学的基盤
生体が透明性を獲得するための第一段階は、光を吸収または反射する要素を発生の過程で完全に排除することである。魚類の体色や不透明性は、主に黒色素胞による光の吸収と、光反射細胞(主にグアニン結晶を含む)による光の反射・散乱に起因する。トランスルーセントグラスキャットは、頭部の一部や内臓を包む腹膜周辺にわずかな銀色や黒色の色素を残すのみで、体幹部においてこれらの色素細胞を完全に欠如している。
ゲノム解析による色素欠損メカニズムの解明
近年の高精度な次世代シーケンシング技術により、この透明な表現型の遺伝的背景が明らかにされている。特筆すべき遺伝子変異として、白皮症に関連するとされるチロシナーゼ関連タンパク質1をコードする推定上の tyrp1b 遺伝子に未成熟終止コドンが発見された。これにより、この遺伝子は機能を喪失した偽遺伝子となっていることが判明した。
さらに決定的な要因として、edn3b 遺伝子が完全に欠失していることが特定された。この遺伝子の役割を検証するために作製されたノックアウト変異ゼブラフィッシュの実験モデルでは、野生型と比較して体表のストライプにおける黒色色素が著しく減少する表現型が確認されている。これらの知見は、edn3b 遺伝子の進化的な喪失が、本種の透明な表現型の獲得に直接的に寄与していることを証明している。
皮膚層のメタマテリアル構造
内部の組織が透明であっても、外界と接する境界である皮膚が不透明であれば、透明性は成立しない。一般的な魚類は物理的保護のためにウロコを持つが、トランスルーセントグラスキャットをはじめとする透明なナマズの仲間は、ウロコを完全に欠如している。
この無ウロコの皮膚は、平均厚さが約20 μmという極めて薄い構造を有している。電子顕微鏡による微細構造解析によれば、この皮膚は主に直径30 nmから40 nmの微細なコラーゲン原線維によって構成されている。これらのコラーゲン原線維は、厚さ80 nmから400 nmの層を形成し、隣り合う層が互いにほぼ垂直に交差する「合板状(plywood structure)」に積み重なっている。このようなナノスケールでの規則的な配向は、可視光領域における散乱を相殺し、光学的に損失のない保護フィルムとして機能している。
| 組織階層構造的特徴 | 寸法 | 光学的機能と効果 |
|---|---|---|
| 体表全体 | ウロコの完全欠如 | 光の巨視的な吸収と表面反射の防止 |
| 皮膚層 | 厚さ約20 μm。直交合板状コラーゲン線維 | コヒーレント散乱の抑制。90%以上の高透過率 |
| 細胞膜レベル | 屈折率の平滑化 | 組織境界でのレイリー散乱およびミー散乱の最小化 |
筋肉組織における透過型回折と動的構造色
生物界において観察される構造色の大部分は、モルフォチョウの羽や甲虫の殻に見られるように「反射」によって生じる。しかし、トランスルーセントグラスキャットの体全体に見られる虹色の輝きは、透過によってのみ存在する極めて稀な光学的現象である。この虹色の根源は、筋原線維内のサルコメア(筋節)が形成する周期的なバンド構造にある。
ミオシンの六方細密充填格子による回折
サルコメアは骨格筋の収縮単位であり、アクチンとミオシンが極めて規則正しく配列している。透過型電子顕微鏡での観察によれば、ミオシンフィラメントが横断面において高度に秩序化された六方細密充填格子(Hexagonal lattice)を形成していることが明らかになった。この横方向のナノスケールでの均一性が、光路内における不規則な散乱を強力に抑制している。
一方、長軸方向においては、電子密度の異なるタンパク質群が周期的に並ぶことで、屈折率の周期的な変動が生じる。この構造が、自然界における高精度の「透過型回折格子(Transmission grating)」として機能し、生体内を通過する光を波長ごとに回折させるのである。生きた魚に見られる虹色の光沢は、主に皮膚に近いサルコメアの周期構造(約2.0 μm)による回折光が支配的である。
生体構造の崩壊と白濁化のメカニズム
生時にはガラスのように透き通り、虹色の回折光を放つ魚体が、死を迎えた直後、あるいは極度のストレスに晒されると、瞬く間に乳白色へと変貌する。この現象は、細胞レベルの光学的な不均一性の増大、すなわち光散乱の急増に起因する。
ATP枯渇とプロテアーゼの活性化
細胞内では、ATP(アデノシン三リン酸)がイオンポンプの駆動や、筋収縮システムのリセットに不可欠である。死後、ATP合成が停止すると、アクチンに結合したミオシン頭部が解離できなくなり、死後硬直が始まる。さらに、イオンポンプの機能不全によりカルシウムイオンが細胞質内に異常流入し、内因性のタンパク質分解酵素であるカルパインを強力に活性化させる。
これらのプロテアーゼ群は、筋原線維の構造完全性を維持するための重要な細胞骨格タンパク質(ジストロフィンやデスミンなど)を無差別に切断・分解する。これにより、筋肉線維の断裂や筋組織の超微細構造が根底から破壊される。
| 生化学的プロセス | 標的・作用メカニズム | 光学的結果(不透明化への寄与) |
|---|---|---|
| ATPの枯渇 | ミオシン頭部の固定、イオンポンプの停止 | サルコメア周期の固定化、動的構造色の喪失 |
| カルパインの活性化 | カルシウム依存性。Z線周辺タンパク質の分解 | 格子構造の破壊による回折パターンの消失 |
| カテプシンの放出 | pH低下に伴う広範な筋肉タンパク質分解 | タンパク質断片の凝集による屈折率の不均一化 |
| 酸化ストレスの増大 | 活性酸素の蓄積、タンパク質のカルボニル化 | 親水性の変化、保水力の低下に伴う細胞間隙液の浸出 |
透過性維持におけるホメオスタシスと進化的トレードオフ
生体が透明性を維持するためには、物理法則(エントロピーの増大)に抗って、筋肉の結晶格子を絶えず能動的に維持しなければならない。これは、生物学的なエネルギー予算において莫大な維持コストを要求する。細胞内の水分量とイオンバランスを制御するイオンポンプの駆動は、細胞の全エネルギー消費の約20%以上を占めるとされている。
透明性と運動性能の二律背反
さらに興味深い事実は、透明性を持たせるための構造的適応が、魚の「運動性能」に対して深刻な進化的トレードオフをもたらしている点である。透明な種は、光の散乱の原因となる筋小胞体と筋原線維の境界面を減らすために、個々の筋原線維を太く進化させた。これにより透過率は飛躍的に向上したが、副産物としてカルシウムイオンの拡散時間が大幅に増大してしまった。
結果として、透明な筋肉は素早い収縮と弛緩を連続して行うことができず、機敏な動きが極端に制限される。本種が捕食者から素早く逃げるという能力を犠牲にして、完全に透明になって隠れるための迷彩に進化のパラメータを振り分けた結果と言えるだろう。
生体模倣と次世代産業技術への応用
トランスルーセントグラスキャットが示すメカニズムは、材料工学や医療分野に対し、革新的なインスピレーションを提供している。
- スマートウィンドウ: 魚の筋肉が白濁するプロセスを応用した、透過率や色を電場で動的に制御できるスマートガラスの開発。
- 人工筋肉とソフトロボティクス: 筋肉の収縮に伴う回折パターンの変化をセンサーとして利用する自己受容感覚を持つロボットの開発。
- 医療用光学イメージング: 特定の分子を用いて生体内の屈折率を均一化し、生きたまま組織深部を非侵襲で可視化する技術。
結論
トランスルーセントグラスキャットが獲得した「究極の透過性」は、光学的、解剖学的、そして遺伝学的な要素が奇跡的なバランスで融合した生体工学の極致である。その透明性は、単なる構造物ではなく、細胞のホメオスタシスによって絶え間なく膨大なエネルギーを供給され、かろうじて維持されている動的な平衡状態に過ぎない。
生命が織りなすこの光学的な魔法の深層を解き明かすことは、単なる基礎生物学の探求にとどまらず、未来のスマートマテリアル、医療デバイス、そして環境調和型テクノロジーの設計思想を根本から刷新する可能性を確固として提示している。



- A chromosome-level genome assembly of the translucent glass catfish (Kryptopterus vitreolus) He, J., et al. (2023). G3: Genes, Genomes, Genetics. (edn3b遺伝子の欠損と透明化表現型の相関に関する解析)
- Transmission-based structural coloration in the glass catfish Cai, X., et al. (2023). Proceedings of the National Academy of Sciences (PNAS). (サルコメアによる透過型回折格子の物理メカニズム)
- Biophysical principles of transparency in freshwater fish species Journal of Experimental Biology. (屈折率マッチングとコラーゲン層のナノ構造に関する研究)
- Evolutionary trade-offs between visual camouflage and swimming performance in transparent fishes Gunderson, A. R., et al. (2022). Biological Reviews. (透明性と運動能力の進化的トレードオフ)
- Post-mortem changes in the optical properties of fish muscle tissue International Journal of Food Science & Technology. (ATP枯渇に伴う白濁化と散乱現象の生化学的解析)
- Tunable smart windows based on smectic A liquid crystal composites Advanced Functional Materials. (生体模倣による動的透過制御材料の応用)





コメント