

魚類における味覚と嗅覚は、人間とは異なる独自の進化を遂げてきました。「水」という共通の媒体の中で、魚たちはどのようにして「味」を感じ、餌を選別しているのでしょうか。本記事では、魚類が甘味を感じない理由や、全身が舌のような構造を持つナマズの驚異的な能力、そしてデンプン質の餌に対する反応の真実について、最新の生理学と分子生物学の視点から解き明かします。
1. 序論:水棲環境における化学受容の特異性と進化
1.1 水中における「味」と「匂い」の境界
脊椎動物の感覚システムにおいて、化学受容は環境情報を取得するための最も原始的かつ基本的な手段です。陸生脊椎動物、特に人間において、味覚と嗅覚は、受容される化学物質の物理的状態によって明確に区別されます。すなわち、嗅覚は空気中を漂う揮発性分子を捉える「遠隔化学受容」であり、味覚は口腔内の液体に溶解した水溶性分子を捉える「接触化学受容」です。
しかし、魚類が生息する水環境においては、すべての化学刺激物質は水という溶媒に溶解した状態で存在します。物理的な運搬媒体が同一であるため、魚類における味覚と嗅覚の区別は、刺激物質の物理的性状ではなく、解剖学的な受容経路と神経投射先によって厳密に定義されます。
- 嗅覚:鼻腔内の嗅上皮にある受容細胞が受容し、嗅神経(第I脳神経)を経て終脳の嗅球へ投射される。
- 味覚:味蕾にある味細胞が受容し、顔面神経(第VII脳神経)、舌咽神経(第IX脳神経)、および迷走神経(第X脳神経)を経て延髄の味覚中枢(顔面葉および迷走葉)へ投射される。
1.2 魚類味覚研究の重要性
魚類の味覚は、単なる餌の識別にとどまらず、嚥下(飲み込み)か吐き出し(拒絶)の最終決定を行う生命維持に直結するシステムです。近年、ゲノム解析技術の進歩により、魚類の味覚受容体遺伝子(T1Rs, T2Rs)の多様性が明らかになりつつあります。これらは、魚種ごとの食性や生息環境に応じて劇的な進化的適応を遂げており、人間の味覚モデルをそのまま当てはめることはできないことが示唆されています。
2. 魚類味覚器の解剖学的・神経生理学的特徴
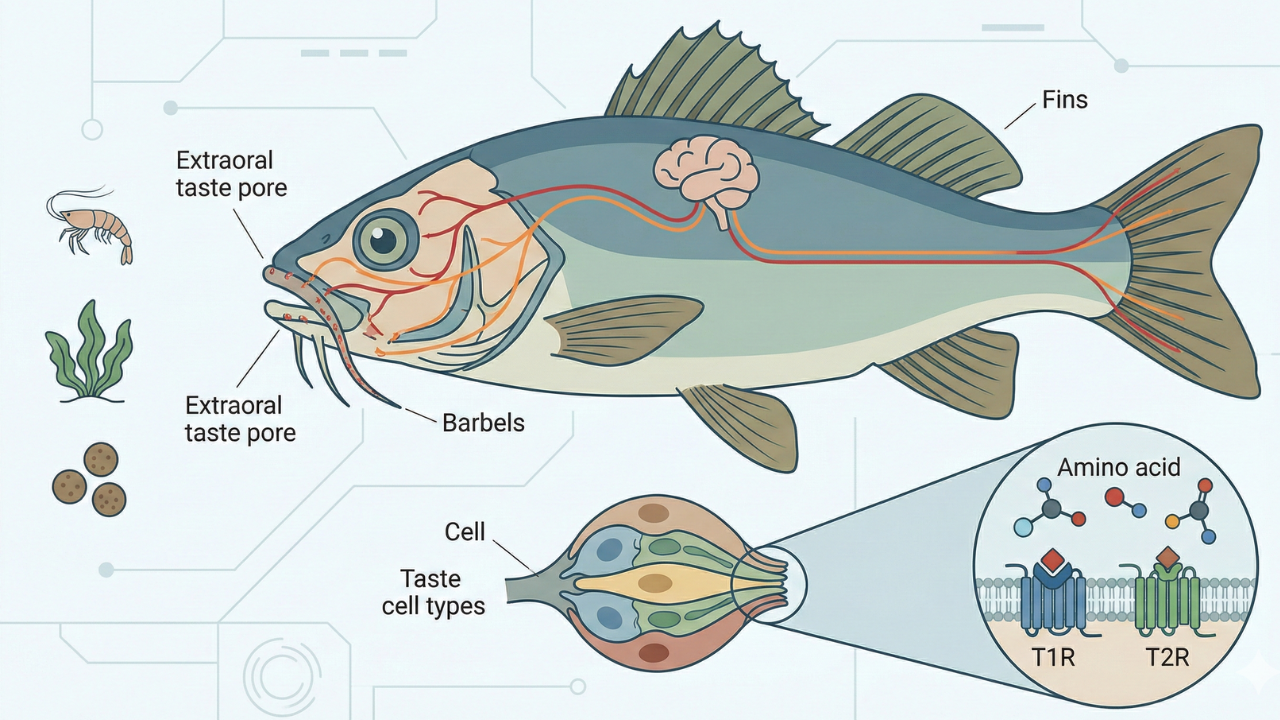
2.1 味蕾の構造と分布:口腔内と口腔外の二重システム
魚類の味覚システム最大の特徴は、味蕾が口腔内だけでなく体表にも分布する点にあります。これは陸生脊椎動物には見られない特徴であり、魚類が環境中の化学情報を全身で受容していることを意味します。
2.1.1 味蕾の微細構造
魚類の味蕾は、タマネギ状またはフラスコ状の微細構造を持ち、主に以下の3種類の細胞で構成されています。
- 暗調細胞:支持細胞としての役割を担うとされる。
- 明調細胞:感覚受容細胞であり、微絨毛を感覚孔に突出させ、水中の化学物質と接触する。
- 基底細胞:細胞の更新や神経伝達の調整に関与する。
2.1.2 口腔外味蕾(体表の味覚)
多くの魚類、特にナマズ目やコイ科の一部、タラ目などは、口唇、口ヒゲ、頭部、さらには体幹部やヒレにまで多数の味蕾を有しています。 特筆すべきはアメリカナマズ(チャネルキャットフィッシュ)であり、彼らは「泳ぐ舌」と称されるほど、全身が味蕾で覆われています。例えば、体長15cm程度の小型のナマズであっても、その体表には25万個以上の味蕾が存在すると推定されており、これは人間の味蕾数(約5,000〜10,000個)を遥かに凌駕します。
この口腔外味蕾は、主に顔面神経によって支配されており、その情報は延髄の顔面葉に投射されます。このシステムは、視界の悪い濁った水中や夜間において、餌の探索と定位に特化した機能を果たしています。ナマズが餌に触れるだけで即座に振り返り、噛み付くことができるのは、この全身に広がる味覚センサーのおかげです。
2.1.3 口腔内味蕾
口腔内、咽頭、鰓弓、および食道入口に分布する味蕾は、舌咽神経および迷走神経によって支配されています。これらの情報は延髄の迷走葉に投射されます。
この口腔内システムは、口に取り込んだ物体が食物であるか異物であるかを識別し、嚥下するか吐き出すかを決定する反射弓を形成しています。特に、底砂ごと餌を吸い込み、口の中で餌だけを選り分けるコイやフナなどの魚種では、迷走葉が巨大に発達しており、口蓋筋系と連携して高度な選別行動を可能にしています。
2.2 神経生理学的応答と行動の乖離
電気生理学的研究(神経からの電気信号の記録)において、味覚神経が特定の物質に応答したとしても、それが必ずしも「好ましい味」として摂食行動を引き起こすとは限らない点に注意が必要です。
例えば、ある種のアミノ酸に対して顔面神経が強い応答を示しても、行動実験ではその物質を摂取しない、あるいは忌避する場合があリます。過去の研究によれば、アラニン、グリシン、プロリンなどは多くの魚種で味覚受容体を強く活性化させますが、これが必ずしも「摂食刺激」として機能するわけではなく、単なる「感知」にとどまる場合もあります。したがって、魚類の味覚を理解するためには、神経応答(感覚入力)と行動応答(運動出力)の両面からのアプローチが不可欠です。
3. 分子レベルで見る魚類の味覚:遺伝子と受容体
脊椎動物の味覚受容には、Gタンパク質共役型受容体(GPCR)であるT1Rファミリー(甘味・うま味)とT2Rファミリー(苦味)が中心的な役割を果たしています。魚類におけるこれらの遺伝子の構成と機能は、哺乳類とは大きく異なり、水中生活と特定の食性に高度に適応しています。
3.1 T1Rファミリー:甘味とうま味の境界の消失
哺乳類において、T1Rファミリーは「T1R1 + T1R3(うま味)」と「T1R2 + T1R3(甘味)」という組み合わせで機能します。しかし、魚類においては、この「甘味受容体(T1R2)」の機能と役割が劇的に変化しています。
3.1.1 魚類のT1R2は「アミノ酸」を好む
遺伝子解析と機能解析の結果、ゼブラフィッシュ(小型熱帯魚)やメダカなどの魚類が持つT1R2/T1R3受容体は、哺乳類が甘味として感じる「糖(スクロースやグルコース)」には反応せず、代わりに「アミノ酸」に反応することが明らかになっています。
具体的には、魚類特有のサブタイプは、プロリン、アラニン、セリンなどのアミノ酸に対して高い親和性を示します。これは、水中環境において糖質が単体で高濃度に存在することは稀であり、一方でアミノ酸はあらゆる生物(餌)に普遍的に含まれるシグナルであるため、進化の過程で受容体のターゲットがアミノ酸へとシフトしたと考えられています。
3.1.2 甘味受容の喪失と保持
一部の魚種、特に肉食性の強い種では、T1R2遺伝子が偽遺伝子化し、機能を失っている例もあります。例えば、ネコが甘味を感じないのと同様に、純粋な肉食魚にとっても糖の検知は生存上の重要性が低いためです。魚類全体として「糖の甘味」よりも「アミノ酸の味(うま味・甘味)」が支配的であると言えます。
3.2 T2Rファミリー:苦味による防御本能
苦味受容体であるT2Rは、生物にとって有害な物質(毒物、アルカロイドなど)を検知し、摂取を回避するための警告系です。デナトニウム(苦味標準物質)やキニーネなどの苦味物質に対し、魚類は明確な忌避行動を示します。
4. デンプンとアミラーゼ:「甘み」を感じる能力の有無
「人間ならデンプン質のものは舌で分解され甘みを感じるが、魚にもそういった能力はあるのか?」という疑問は、魚類の消化生理学の核心に触れるテーマです。結論から言えば、魚類はそのような能力を持っていない可能性が極めて高いと言えます。
4.1 人間における「デンプンの甘み」のメカニズム
人間がパンやご飯を咀嚼すると甘みを感じるのは、唾液中に含まれる唾液アミラーゼの働きによるものです。口腔内で物理的な咀嚼と同時にアミラーゼがデンプン(多糖類)に作用し、マルトース(二糖類)やグルコース(単糖類)へと分解します。これらが舌の甘味受容体を刺激することで、即座に甘味として知覚されます。
4.2 魚類における唾液腺の欠如とアミラーゼ活性
対照的に、魚類の口腔生理学は人間とは根本的に異なります。
- 唾液腺が存在しない:魚類には人間のような唾液腺が存在しません。口腔内の粘液細胞から分泌される粘液が潤滑剤の役割を果たしているに過ぎません。
- 咀嚼行動の欠如:多くの魚類は酵素による化学的消化を行うための滞留時間がほとんどありません。餌は破砕された直後に食道へと送り込まれるため、口腔内でデンプンが糖に分解される時間的猶予が存在しないのです。
4.3 結論:魚はデンプンの甘味を感じない
魚がデンプン質の餌(パンや練り餌)を口にした際、唾液アミラーゼによる分解が起きないため、糖の生成も起こらず、したがって「甘み」を感じることはありません。彼らがデンプン質の餌を好む場合、それはデンプンそのものではなく、添加されたアミノ酸(グルテンや魚粉由来)の味、あるいは匂い、または物理的な食感(テクスチャー)に反応していると考えられます。
5. 「塩味」と「甘味」に対する魚類の具体的反応
5.1 塩味:味覚か、生理的シグナルか
哺乳類が塩味を感じるための受容体(ENaC)は、魚類のゲノムでは欠損しているか機能していません。しかし、魚類は塩分濃度の変化を敏感に察知します。
- 忌避反応:高濃度の塩分に対しては、明確な忌避行動を示します。これは高浸透圧環境が危険であるための回避行動です。
- 誘引・嗜好性:一方で、淡水魚の飼料に1〜2%程度の塩を添加すると、成長や飼料効率が向上する事例があります。これは味覚的な「美味しさ」というよりは、生理的な浸透圧調節のための補償行動として解釈されます。
5.2 甘味:アミノ酸こそが「甘み」
前述の通り、魚類にとっての「甘味」は糖ではなくアミノ酸によって引き起こされる感覚である可能性が高いです。人間が甘みを感じるアミノ酸(グリシン、アラニン、プロリン)は、魚類にとって最強の摂食刺激物質です。魚にとっての「スイーツ」とは、砂糖菓子ではなく、グリシンたっぷりのエビの切り身なのかもしれません。
6. 食性による味覚選好性の比較
6.1 肉食性魚類(ブリ、マグロ、ウナギなど)
アミノ酸への感受性が極めて高く、特にプロリン、アラニン、グリシンなどに強い反応を示します。また、死後分解が進んだ筋肉組織から生じるイノシン酸などの核酸関連物質にも強く反応します。一方で、糖類にはほとんど反応しません。
6.2 草食性・雑食性魚類(ソウギョ、コイ、ティラピアなど)
肉食魚同様にアミノ酸には反応しますが、植物性タンパク質に多い酸性アミノ酸(グルタミン酸など)を好む傾向があります。ティラピアなどはクエン酸などの有機酸にも嗜好性を示します。糖類については、肉食魚に比べれば許容性は高いものの、アミノ酸への反応の方が支配的です。
6.3 果実食性魚類の謎(パクーなど)
アマゾンの水没林で果実を食べるパクー(ミロソマ類)でさえ、実験では砂糖に対する明確な選好性が確認されていません。彼らが果実を見つける手がかりは、着水音(聴覚)、匂い(嗅覚)、色(視覚)であり、口に入れた後の「甘味」は決定的な要因ではない可能性があります。
7. 消化管における「第二の味覚」:腸管味覚
近年の重要な発見として、味覚受容体が口腔内だけでなく、消化管(腸)にも発現していることが挙げられます。これを「腸管味覚(Gut Sensing)」と呼びます。
魚が餌を飲み込んだ後、腸内で消化が進み、アミノ酸やグルコースが遊離すると、腸壁の受容体がこれを感知します。これにより、消化酵素の分泌タイミングを調節したり、脳に「満腹シグナル」を送ったりします。口で「甘み」を感じなくても、腸でグルコースを検知することで、効率的なエネルギー蓄積を行うシステムが備わっているのです。
補遺:主要データ表
| 食性カテゴリー | 代表魚種 | 最も強力な刺激物質 | 糖類への反応 | 備考 |
|---|---|---|---|---|
| 肉食性 | ブリ マダイ ニジマス |
イノシン酸、プロリン アラニン、ベタイン プロリン、グリシン |
無反応 | 魚肉エキスや核酸関連物質に強く反応。視覚情報も重要だが最終判断は味覚。 |
| 雑食性 | アメリカナマズ ティラピア |
アラニン、アルギニン グルタミン酸、アスパラギン酸 |
無反応 一部反応 |
ナマズは全身の味蕾で超高感度にアミノ酸を検出。ティラピアは有機酸も好む。 |
| 草食性 | ソウギョ | システイン、グルタミン酸 | 条件付きで反応 | 植物成分とアミノ酸の両方に反応する。 |
| 果実食性 | パクー | (研究途上) | 明確な選好性なし | 果実食だが糖への反応は薄い。嗅覚や聴覚への依存度が高い可能性。 |
| 特徴 | 人間 | 魚類 | 影響・結果 |
|---|---|---|---|
| 唾液腺 | 有 | 無(粘液細胞のみ) | 魚は食物を化学的に処理する液体を持たない。 |
| 唾液アミラーゼ | 高活性 | 欠如(または極めて微弱) | 魚の口内ではデンプン分解が進まない。 |
| 口腔内滞留時間 | 長い(咀嚼あり) | 短い(丸呑み) | 魚は反応が起きる前に飲み込んでしまう。 |
| デンプンの味 | 「甘い」 | 「無味」(食感のみ) | 魚はパンの甘みを感じていない可能性大。 |
| グルコース受容 | 舌の受容体で感知 | 主に腸の受容体で感知 | 魚の糖感知は摂食後(代謝)のプロセス。 |








コメント